The leaves change shape with age. The aerial roots are not decoration. The geniculum is a joint, not a hinge. And every fenestration is a story about light. Monstera anatomy is where every Monstera care decision actually comes from.
Why Anatomy Is the Compass
Most Monstera advice tells you what to do. Water when the top two inches are dry. Bright indirect light. Provide a moss pole. The advice isn't wrong, but it gives you no way to predict what the plant will do next or to recognize when conditions have changed.
Anatomy gives you that prediction engine. A Monstera is not a houseplant. It's a climbing hemiepiphyte from a Central American rainforest, and almost every odd thing it does in your living room is a holdover from that life. The juvenile-to-adult leaf transition, the aerial roots reaching for nothing, the strange swollen knuckle where the leaf meets the petiole, the holes that aren't supposed to be there. Each one is a structure built for a job the plant evolved to do.
This essay walks the Monstera body at the resolution that's useful for cultivation. Light navigation, leaf development, the climbing apparatus, the holes, the roots, and the inflorescence. For each, the practical thing that changes about how you grow it. No pure academia, no skipped fundamentals.
A Seedling Walks Toward Shadow
Start with how a Monstera arrives in the world, because the strangest fact about the genus is how it begins.
A Monstera seed lands on the forest floor in the deep shade of a Central American rainforest. When it germinates, the seedling does something almost no other plant does: it grows toward the darkest patch of horizon it can see. Not toward light. Toward shadow.
The behavior is called skototropism, from the Greek skotos for darkness, and it was documented by Donald Strong and Thomas Ray at Harvard in the mid-1970s working with Monstera gigantea in Costa Rica. They placed seedlings near black cardboard targets and watched them grow toward the targets in a way that light-loving plants never do. The targets were proxies for tree trunks. In a forest, the darkest object on the horizon is the trunk of the largest tree, and the largest tree is the best climbing host. The seedling is hunting for something to climb.

Once a seedling reaches a trunk, the rule flips. The plant switches from skototropism to ordinary phototropism, starts climbing upward into the brighter canopy, and begins producing the larger, more divided leaves that define the adult form. Until then, juvenile Monstera growth looks almost nothing like the plant you bought.
You'll see the residue of this behavior in your own collection. A Monstera in a too-dim corner sometimes sends long, thin runners along the floor or across a shelf with small, undeveloped leaves. That's not stretch growth in the usual sense. That's a hemiepiphyte still looking for a host tree, still in skototropic mode, still trying to find the trunk it never gets to find. Give it something vertical to climb and the runners stop. The plant transitions.
Heteroblasty: The Plant That Changes Its Mind
The juvenile-to-adult transition has a name: heteroblasty. It refers to a plant that produces structurally different leaves at different life stages, not gradually but in distinct phases. Monstera is one of the most studied heteroblastic plants in botany, in large part because of Thomas Ray's work in the 1980s and 1990s on leaf morphogenesis in the genus.
Juvenile Monstera leaves are small, heart-shaped, entire (no holes, no divisions), and pressed close to whatever the plant is climbing. The growth habit is called shingle: each leaf flattens against the bark in a roof-tile arrangement. Monstera dubia is the most extreme example. As a juvenile it produces tightly-shingled, dark-green, almost reptilian foliage against the trunk. The adult form of the same plant, higher up in the canopy, looks like an entirely different species: large, deeply divided, dramatically fenestrated leaves on long petioles that stand away from the host.

For most Monstera in cultivation, including M. deliciosa and M. adansonii, you see a compressed version of this transition. Small unfenestrated juvenile leaves. Then medium leaves with a few holes. Then larger leaves with more holes and the first lobes. Then the full adult form. The shift is driven by a combination of plant size, light availability, and the presence of a climbing surface. A Monstera that never gets to climb often never produces fully adult leaves, no matter how old it is.
Practical consequence: if you want the dramatic adult foliage, the plant needs to climb. A moss pole or coir totem isn't a stylistic choice. It's a developmental trigger. The plant reads vertical attachment as "I have reached the tree" and shifts its leaf-production program accordingly. Without that signal, you get a juvenile plant in a large pot, which is what most apartment Monsteras are.
The Climber's Architecture
Monstera is not a vine. It's a hemiepiphyte, a category botanists use for plants that begin life on the ground and end it in the canopy, or begin in the canopy and send roots to the ground. Monstera does the first version. It germinates terrestrially, climbs a host tree using aerial roots, and once established may eventually lose its connection to the soil entirely, surviving as a canopy plant rooted into bark crevices and the host tree's own decomposing debris.
The architecture is built for this two-stage life. The stem is thick, soft, and persistent. Internodes are short on juvenile growth, where the shingle pressed against the trunk needs to grip tightly, and longer on adult growth, where the canopy leaves can spread out into space. The petioles get longer with age and the leaves get bigger, because the adult plant lives in a light environment the juvenile never experienced.
None of this works without the aerial roots, which deserve their own section. But the structural takeaway is that a Monstera in a pot is, in a meaningful botanical sense, only half a plant. The other half is the tree it's supposed to be growing on. Everything you do for the plant in cultivation is, one way or another, an attempt to substitute for that missing tree.

The Geniculum: Joint, Not Hinge
Look at the petiole of a mature Monstera leaf and you'll see a small, slightly swollen segment where the petiole meets the leaf blade. It looks like a tiny knee. That's the geniculum, from the Latin for "little knee," and it's one of the most diagnostic features of the genus.

It is not just thicker tissue. The geniculum is a true motor organ, anatomically analogous to a pulvinus: a cluster of specialized parenchyma cells that can change turgor pressure rapidly and re-orient the leaf blade. The blade above the geniculum tilts to track light, to shed water after rain, and to reorient when the plant is repositioned. The movement is slow on a houseplant timescale, hours rather than seconds, but it's real, and it happens whether you notice it or not.
Some growers mistake the geniculum for a weak point and try to support it. That's exactly wrong. The geniculum is structural, not damaged. If it's swelling, browning, or collapsing on a particular leaf, the plant is telling you something about hydration or temperature, not about its skeleton. A healthy geniculum looks like a slightly enlarged section of petiole, slightly paler than the rest, sometimes with a faint wrinkle on the underside.
Practical consequence: when you rotate a Monstera, give it a few days before judging the new orientation. The geniculum is repositioning the leaves to the new light angle, and the visible result lags the input by a noticeable margin. The plant is doing slow physics on every leaf, all the time.
Fenestration: A Story About Light
Now the holes.
The fenestrations of Monstera deliciosa and the splits of M. adansonii are among the most recognizable leaf structures in the plant kingdom, and they are also the source of more bad theories than almost any other feature in the genus. Three hypotheses have circulated for a century: that the holes let wind through and reduce mechanical damage in storms; that they shed water during heavy rain; and that they let dappled light reach lower leaves on the same plant. All three have intuitive appeal. Only one of them has held up.
The cleanest test came from Christopher Muir in a 2013 paper in The American Naturalist. Muir modeled the light environment of a Monstera leaf in the rainforest understory, where most of the available photosynthetic energy arrives as brief sunflecks: shafts of direct sun that punch through gaps in the canopy and move across the forest floor as the day goes on. A leaf with no fenestrations catches every sunfleck that hits it, but covers a small footprint. A fenestrated leaf with the same total leaf area covers a much larger footprint, intercepting sunflecks from a wider patch of horizon. The total light captured per leaf rises, even though the holes themselves catch nothing.

The strategy makes sense only in a deeply shaded understory where sunflecks are the dominant source of usable light. In open sun, the same architecture would be a waste of leaf. That's why fenestrations show up in shade-dwelling climbing aroids again and again (Monstera, Rhaphidophora, some Epipremnum) and almost never in plants that live in continuous bright sun.
It also explains the heteroblastic pattern. Juvenile Monstera leaves, growing low on the trunk in deep shade and pressed against bark, don't fenestrate. There are no sunflecks to chase down there. Adult leaves, growing higher in the canopy where sunflecks are abundant, fenestrate heavily. The plant only makes holes when holes pay off.
Practical consequence: if your Monstera is producing small, unfenestrated leaves indoors, the problem is almost never humidity or fertilizer. It's that the plant is reading its position as "still juvenile, still on the forest floor." Climbing surface and light level are the levers. A mature plant on a tall moss pole in a south-facing window with diffused light will fenestrate readily; the same plant in a low pot in a dim corner will produce shingle-like juveniles for years.
The Aerial Root
If you've kept a Monstera for any length of time, you've watched aerial roots emerge from the stem and reach out into the room. They are easily the most-asked-about feature of the plant. The common questions: should I cut them, should I bury them, should I let them dangle, are they normal.
The answers come from understanding what the root is for.
Monstera aerial roots are dimorphic: they come in two functional types, sometimes from the same node. Anchor roots are short, stiff, often slightly flattened, and they emerge near the climbing surface. Their job is mechanical attachment. In the wild they wedge into bark crevices and grip the host tree. In your living room they tend to push toward the nearest wall or moss pole and try to do the same thing. Feeder roots are longer, thicker, and more flexible. They descend toward the ground or toward any consistent source of moisture and nutrients, including the substrate in the pot. In the canopy they can reach many meters down a host tree to tap leaf litter at the base.
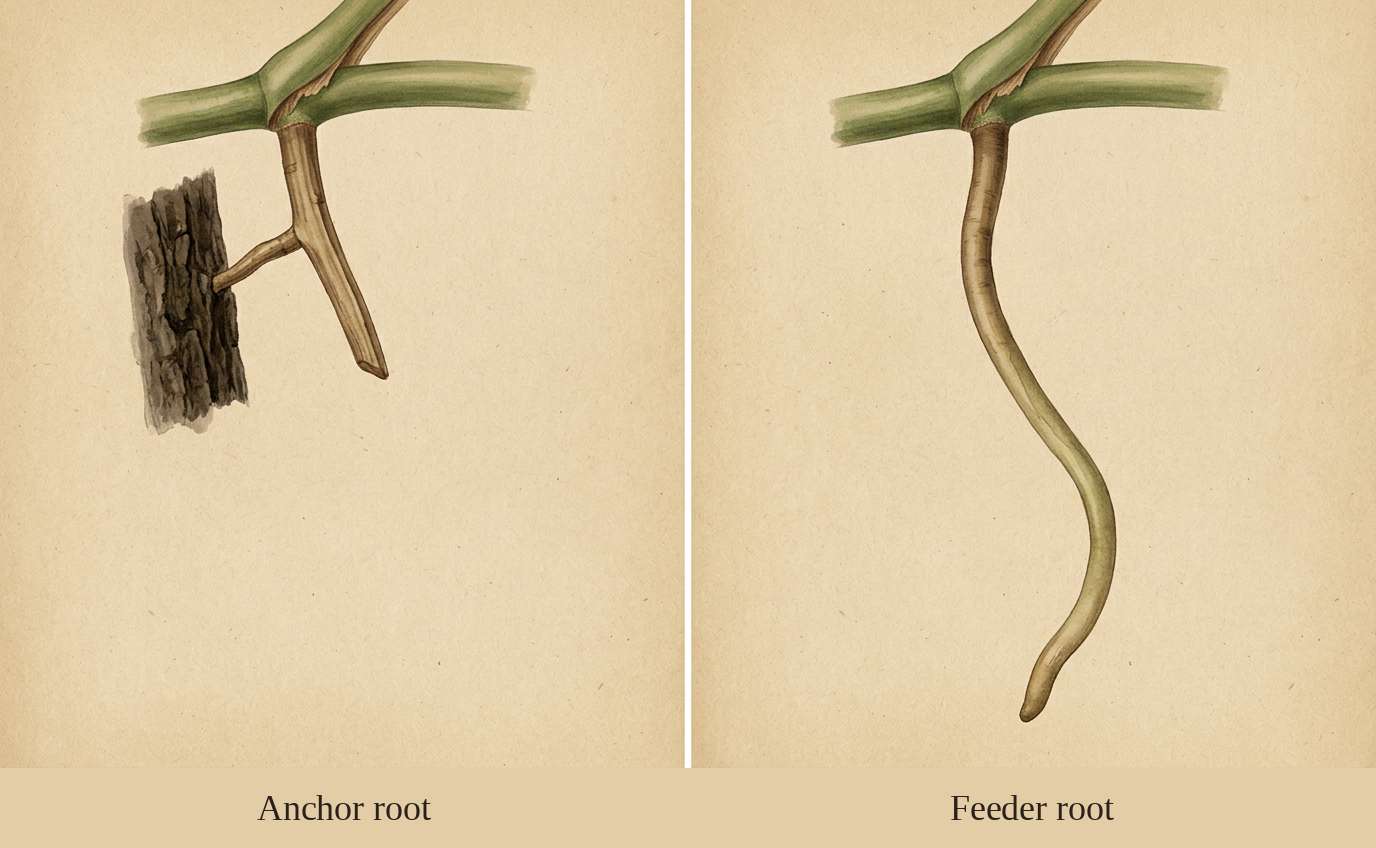
Both types are functional. Cutting them won't kill the plant, but it does remove tissue the plant invested energy in producing. If an anchor root has already attached to a moss pole or a wall, leave it. If a feeder root is reaching the ground or your substrate surface, leave it. The only aerial roots worth removing are ones that have died back (brown, hollow, dry) or ones that are growing into spaces where they will physically damage furniture or walls.
One persistent myth: aerial roots are not primarily photosynthetic, and they are not pulling significant moisture from the air. In a forest with 90% relative humidity that distinction matters less, but in a 40% living room the air-absorption pathway contributes almost nothing. The roots want either substrate (for water and nutrients) or solid attachment (for anchoring). Misting them is mostly theater.
The Thai Constellation in my living room (south-east window with supplemental grow-light) is climbing a cedar plank, but its aerial roots are doing something separate. Two of them have grown roughly 18 inches (46 cm) sideways across the wall in opposite directions, clearly looking for something else to attach to. I haven't pruned them because they're working exactly the way they would in a Costa Rican forest, except the host tree is my drywall.
The plant's adult leaves arrived about four months after I gave it the cedar plank. Same plant in the same room, just given vertical attachment, produced its first deeply fenestrated leaf within a single growth cycle. The geniculum on those new leaves is noticeably more developed than on the juvenile growth lower down. Anatomy reads attachment as a developmental signal, and the signal is unmistakable.
The Inflorescence and the Fruit
Most growers will never see a Monstera flower indoors, but the flower is worth understanding because it tells you what the plant is botanically and because M. deliciosa produces a real, edible fruit that almost no other houseplant matches.
The inflorescence is a classic aroid structure: a fleshy central column called the spadix, surrounded by a single creamy-white modified leaf called the spathe. Anthurium and Spathiphyllum have the same architecture in different proportions. In Monstera the spathe is hooded and the spadix is dense with tiny flowers covering its entire surface.

After pollination, the spadix matures into the fruit. The fruit of M. deliciosa ripens slowly, over roughly twelve months, and when fully ripe (the hexagonal scales pop off from the base upward) the flesh is famously pineapple-banana in flavor. Hence "deliciosa."
The catch is that the fruit is dangerous before it's ripe. Like most aroids, Monstera tissues contain microscopic needles of calcium oxalate called raphides, packaged in specialized cells called idioblasts. Bite into unripe Monstera fruit and the raphides eject into your mouth and throat, causing severe irritation. The same chemistry is why every Monstera leaf is mildly toxic to pets and small children. The plant isn't being aggressive; it's defending its tissues with a strategy shared across the entire Araceae family.
Practical consequence: don't worry about touching the plant. Calcium oxalate raphides need to be ejected from cells, which means the tissue has to be bitten or chewed. Routine handling is fine. Keep the unripe fruit and any chewed leaves away from kids and pets, and treat sap from cuts the way you'd treat any irritant on bare skin.
The Takeaway
Monstera is a climbing hemiepiphyte from a Central American rainforest. Its body is built for two phases: a shaded juvenile on a forest floor, hunting for a tree trunk to grip, and a sunfleck-chasing adult in the canopy with large, fenestrated, light-tracking leaves. Every anatomical structure on the plant has a job in one phase or the other. Aerial roots anchor it and feed it. The geniculum re-orients the blade to catch light. Fenestrations widen the leaf's footprint without growing more tissue. Heteroblasty lets the plant produce two different leaf forms for two different environments. Skototropism gets it to the tree in the first place.
Almost every care decision worth making comes back to one of those facts. The collector who reads the leaves as a sunfleck strategy will use a moss pole differently from the one who thinks of fenestrations as decoration. The collector who knows the aerial roots are dimorphic will stop cutting them off. The collector who recognizes the geniculum as a working joint will not panic when the plant repositions its leaves overnight. The science isn't separate from the practice. It's the practice, just one layer down.
Madison, M. (1977). A revision of Monstera (Araceae). Contributions from the Gray Herbarium of Harvard University, 207, 3–100.
Muir, C. D. (2013). How did the Swiss cheese plant get its holes? A test of two hypotheses. The American Naturalist, 181(2), 273–281. https://doi.org/10.1086/668819
Strong, D. R., & Ray, T. S. (1975). Host tree location behavior of a tropical vine (Monstera gigantea) by skototropism. Science, 190(4216), 804–806. https://doi.org/10.1126/science.190.4216.804
Ray, T. S. (1990). Metamorphosis in the Araceae. American Journal of Botany, 77(12), 1599–1609. https://doi.org/10.1002/j.1537-2197.1990.tb11399.x
Mayo, S. J., Bogner, J., & Boyce, P. C. (1997). The Genera of Araceae. Royal Botanic Gardens, Kew.
Putz, F. E., & Holbrook, N. M. (1986). Notes on the natural history of hemiepiphytes. Selbyana, 9(1), 61–69.
Plants of the World Online. Monstera Adans. Royal Botanic Gardens, Kew. https://powo.science.kew.org/